A SPATIAL MODELING AND DECISION SUPPORT SYSTEM FOR CONSERVATION OF BIOLOGICAL DIVERSITY
Wildlife habitat modeling
Many of the issues in biodiversity assessments revolve around a central problem of synthesizing data from many disparate sources, compiled at different times, by different methods, and with different reliability. For rapid assessments of the status of biodiversity, we must utilize these existing sources, but the problem remains of how to best utilize each of them for its strengths and to track areas of uncertainty or ignorance. Recent advances in automated systems that handle spatial data revolutionize the way we can represent distributions of plant and animal species. Traditional dot maps, grids or line drawings used until very recently to show species ranges are being replaced by applications of models to maps of habitat variables (e.g., Walker 1990, Scott et al. 1993). Models of ranges of populations or metapopulations now take explicit account of spatial effects e.g. patch size, distance between suitable habitat patches, and the qualities of contiguous habitats (e.g. Hanski and Gilpin 1991). Models of the behavior and movements of individual organisms now are carried out on spatial grids which accommodate effects such as territoriality and foraging (e.g. Folse 1989). The range of environmental data to support these modelling efforts has also grown exponentially, due to technological advances (i.e. improvements in digitizing, satellite and aircraft remote sensing, telemetry, and Global Positioning Systems).
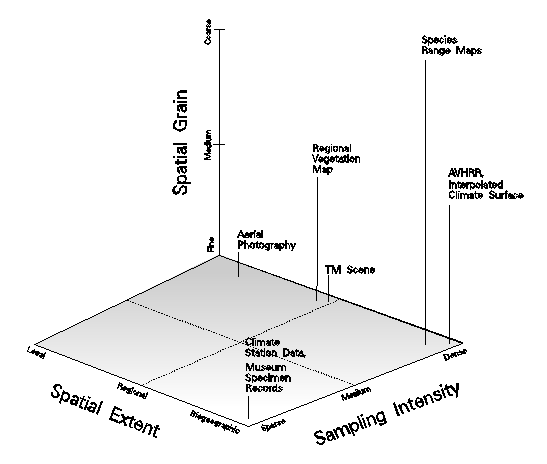
Geographic information systems technology permits the generation of complex representations of species distributions, while most of the data underlying these patterns are coarse. This suggests the importance of structuring such data along axes of differing data extent, tiling schemes, themes, and time, and displaying different representations of distributions, the philosophy being that comparison of multiple representations provides a sense of the actual distribution through convergence of evidence. Our research focused on integration of biodiversity data of different temporal and spatial scales (Figure 10) into more complete representations of species distributions (Stoms et al. 1996). We followed this conceptual approach using a lizard, the orange-throat whiptail (Cnemidophorus hyperythrus), which is native to southern California. The analysis was hierarchically structured by first mapping overall lizard range limits, then suitable habitats within the range, and then habitats over a local extent (Hollander et al. 1994, Figure 11). Data sources include a generalized range outline, museum records, and field observations, as well as climate data, vegetation maps, and satellite imagery to serve as associated environmental variables. Comparison of representations resulting from these different data sources makes biases evident, highlights areas of inadequate sampling, and can lead to new inferences about habitat relationships. The study helped identify many of the obstacles found in commercial GIS packages for doing multiscale data integration or for linking GIS databases with analysis tools used by vegetation ecologists.
Figure 10. Scale and spatial sampling. Three dimensions are shown to characterize the scale of biodiversity-related data sets, these being spatial grain, spatial extent, and sampling intensity. Similar axes could be applied in the temporal domain.
The technology of GIS has greatly enhanced the ability to visualize natural resource data. Digital geographic data are playing an increasing role in wildlife management, providing computerized representations of wildlife habitat, administrative boundaries, significant cultural features, and of particular interest here, wildlife species distributions. GIS has much potential for enhancing the visualization of the latter. The distribution of a species is a complex dynamic phenomenon which is poorly represented by a static map. But complex models of a species' distribution which reflect a variety of environmental factors can be developed in a GIS environment. Unfortunately the complexity of GIS may make it difficult for wildlife managers to take advantage of this potential.
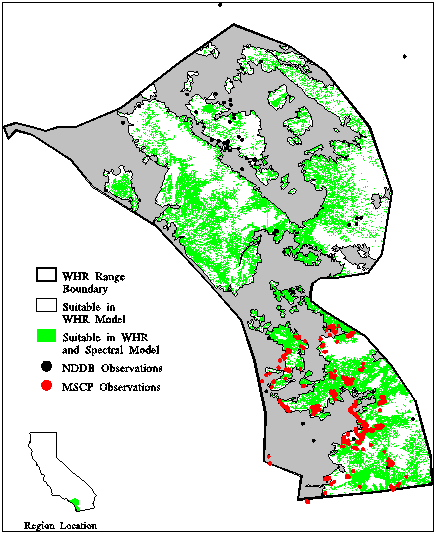
Figure 11. Whiptail distribution within its range limits. Areas not shaded in gray are suitable according to the WHR model; superimposed in green are pixels identified in spectral classification as likely to contain whiptail observations. Also superimposed are point observation data from two data sets.
There are a variety of reasons for these difficulties. Fundamentally these stem from the fact that expert wildlife biologists do not necessarily have the experience with these computer systems to fully utilize their capabilities. Geographic information systems are complex systems, and this complexity is still present even with the best-designed interface. Furthermore, additional work is often needed to integrate specialized datasets into the GIS. Thus it is necessary to give special attention to the process of extracting information from the wildlife expert.
A model predicting the relative abundance of the wild pig at a regional scale was developed for the central coast of California. Components of this model include habitat suitability and human disturbance, specifically using the WHR habitat type, distance from water sources, road density, and hunting pressure as factors in the model. This model used an influence diagram representation to structure the relationship between variables in the model. Interactive review of the model was accomplished by displaying the GIS layers of the model at a workshop of expert biologists and incorporating their comments. Workshop participants approved of the final version of the model, which predicted relatively high pig abundance in Monterey, San Benito, and Santa Clara counties, and low abundance in the Bay Area, Salinas Valley, and around Monterey Bay, with much local-scale variability.
The Relative Abundance layer synthesizes the various GIS layers to produce a map showing the relative abundance of pigs over the ecoregion. This combines both the human disturbance and habitat suitability components. As in the disturbance layer, the degree of hunting pressure is a parameter that is changed in different versions of this model. In the version originally presented to workshop participants, heavy hunting pressure depresses abundance by 50%. In this map, areas with high pig abundance include portions of Monterey, San Benito, and Santa Clara counties. Areas with low pig abundance include the urbanized Bay Area, the Salinas Valley, and areas around Monterey Bay. There is much local-scale variability in this map, reflecting the influence of a number of factors in the component layers - e.g. different distances from water and different road density effects.
Finally, a model was developed predicting pig habitat suitability at a local scale, that is using a 30 meter pixel size across an region about 1000 sq. km. in size. This is a spatialized habitat model - that is, it integrates habitat characteristics over a landscape the size of the home range of the pig. The model incorporates food availability, cover availability, and the degree of access to water. This model was tested for two study regions - one in San Benito county, another in San Luis Obispo county. Variants of the model made use of different values for the home range radius of the pig. Experts at the aforementioned workshop agreed to the variant with a radius of 1 mile from food and cover, and up to 3 miles from water. There was a high degree of correlation between areas that provided food and areas that provided cover; therefore suitability is mainly a function of the interaction between access to woody vegetation and water. Neither the regional or the local model have been adequately validated. Further field work is needed to do this and both models will be refined as more information becomes available on influences on pig habitat and distribution.
This 30-meter scale is quite finely resolved with respect to the home range of a pig. This means that assessment of the potential for each pixel to be utilized by pigs will depend upon integrating the habitat characteristics over the area surrounding the pixel within roughly the home range radius of a pig. This dependence on the landscape context is why this type of model is termed a spatial habitat model. The aim of this model is a map that shows the average utilization of the landscape by pigs over a long period of time.
The main characteristic of this local-scale model is that it integrates the quality of the landscape over the presumed home range of a pig. Thus it views habitat suitability not as a point function but as a function that depends on the surrounding neighborhood. In spatial terms, a smoothing filter with the radius of the home range has been applied across the landscape. Thus many of the layers in the model have a fuzzy appearance to them. One drawback to this smoothing process is that it is computationally intensive: with 30 meter pixels and a home range radius of 1 mile, calculations must be carried out over a circular buffer 53 pixels in radius. This spatial integration is a fairly novel approach in GIS-based wildlife habitat modelling: usually habitat suitability is considered on a pixel-by-pixel basis.
With respect to GIS methodology, this project has illustrated how expert review of the component layers in a GIS model can enhance the modelling process. One recurrent problem with GIS modelling is that the mapped results of the model are presented without showing how these have been derived, though their derivation is needed for their proper interpretation. The methodology used here illustrates how interactive review of the components of the GIS model can aid in its development. This has been accomplished in a workshop where the components of GIS models were presented to wildlife experts. The portability of the presentation has been facilitated by technology such as the use of laptop computers, overhead display devices, and current GIS visualization software. Nevertheless, the present technology did not allow these models to be altered interactively during the workshop. Another issue concerning expert review is whether to elicit feedback at a workshop, as done here, or through interviews with experts carried out individually. The latter offers the possibility of reaching the opinion of more people, whereas group review allows for more synergism and consensus from participants.
From the point of view of wildlife habitat modelling in general, there are a number of results to be highlighted from this study. The first is the method used to generate maps of wildlife habitat types (WHR classes) at a 30-meter pixel resolution. This is carried out through the use of ancillary map information to drive a classification of a Landsat Thematic satellite image. Essentially, the taxonomic detail in the coarse-scale ancillary map is used to refine the coarser classification of the more finely spatially resolved satellite image. Another element to the wildlife habitat modelling is incorporation of human disturbance as a factor affecting relative abundance levels. This is done multiplicatively, with higher levels of hunting pressure or greater road densities corresponding to lower relative abundances. Also, additional influencing factors can be incorporated in the network model by adding nodes and links to the diagram. Finally, both the local and regional models are spatialized in that they integrate habitat factors across the landscape rather than focusing on a single point. This is a step towards building dynamic models of wildlife populations through space and time.