What's New
Research & Projects Publications
People
Data
Links
3. PREDICTED ANIMAL DISTRIBUTIONS AND SPECIES RICHNESS
Introduction
Methods
Results
Accuracy Assessment
Limitations and Discussion
IntroductionAll species range maps are predictions about the occurrence of those species within a particular area (Csuti 1994). Traditionally, the predicted occurrences of most species begin with samples from collections made at individual point locations. Most species range maps are small-scale (e.g., >1:10,000,000) and derived primarily from point data for field guides. The purpose of the Gap analysis vertebrate species maps is to provide more precise predictions of the current distribution of individual native species within their general ranges. With this information, better estimates can be made about the actual amounts of habitat area and the nature of its configuration.
Gap analysis maps are produced at 1:100,000 and are intended for applications at the landscape or "gamma" scale (heterogeneous areas generally covering 1,000 to 1,000,000 hectares and made up of more than one kind of natural community). Applications of these data to site- or stand-level analyses (site - a microhabitat, generally 10 to 100 square meters; stand - a single habitat type, generally 0.1 to 1,000 ha; Whittaker 1977, see also Stoms and Estes 1993) are likely to be compromised by the finer-grained patterns of environmental heterogeneity that are resolved at those levels.
Gap analysis uses the predicted distributions of native vertebrate species to evaluate their conservation status relative to existing land management (Scott et al. 1993). However, the maps of vertebrate species distributions may be used to answer a wide variety of management, planning, and research questions relating to individual species or groups of species.
Previous to this effort there were no maps available, digital or otherwise, showing the likely present-day distribution of species by habitat type across their ranges. Because of this, ordinary species (i.e., those not threatened with extinction or not managed as game animals) are generally not given sufficient consideration in land-use decisions in the context of large geographic regions or in relation to their actual habitats. Their decline because of incremental habitat loss can, and does, result in one threatened or endangered species "surprise" after another. Frequently, the records that do exist for an ordinary species are truncated by state boundaries. Simply creating a consistent spatial framework for storing, retrieving, manipulating, analyzing, and updating the totality of our knowledge about the status of each vertebrate species is one of the most necessary and basic elements for preventing further erosion of biological resources.
The species nomenclature used in this report generally follows the standards of The Nature Conservancy's (TNC) Central Scientific Databases for animals. Where this standard is simply a name change, we have used the new TNC name along with the WHR code number. In a few cases, the reclassification of species has led to a split or aggregation of species in the WHR database. It was often not possible to reasonably model the distribution of such a taxon, and in such cases we have retained the original taxonomy as defined for the WHR database.
Specific taxonomic issues were resolved as follows:
- The Yellow-bellied sapsucker, as currently classified, does not occur in California. Populations formerly assigned to Sphyrapicus varius are now called S. nuchalis, the Red-Naped Sapsucker.
- The Western flycatcher (Empidonax difficilis) has been split into two distinct species, the Pacific-slope flycatcher (which retains the original scientific name) and the Cordilleran flycatcher (E. occidentalis). Because the two species occupy distinct geographic ranges in California, we were able to associate the portions of the overall range with the appropriate species and predict their habitats in this gap analysis. E. occidentalis was assigned a new WHR code for this project (B990).
- A similar situation occurred with the Black-tailed gnatcatcher. Polioptila melanura was split into a desert-dwelling Black-tailed species which retained the scientific name, and the California gnatcatcher (P. californica) in coastal sage habitats. The latter was assigned a WHR code of B991, and the two species were modeled independently in this analysis.
- Shrews along the west coast have been revised. The Pacific shrew (Sorex pacificus) no longer occurs in California. The populations formerly called S. pacificus are now assigned to S. sonomae, the Fog Shrew.
- The Pacific giant salamander (Dicamptodon ensatus) was recently split into three species, two of which occur in California (D. ensatus and D. tenebrosus). We were unable to distinguish the individual ranges of these two species and chose to leave these two species as one for this analysis. Conclusions about the management status of either species may not be reflected accurately in the results for the combination of the two.
- The Olympic salamander was split into two species. Currently, Rhyacotrition olympicus does not occur in California, but the range is occupied by R. variegatus, the Southern Torrent Salamander.
- The slender salamanders are undergoing major revisions in taxonomy. Because of the volatility of the nomenclature and uncertainties about the ranges of new species, we kept the California slender salamander (Batrachoseps attenuatus) and the Pacific slender salamander (B. pacificus) as portrayed in the California Wildlife Habitat Relationships System.
- The Spotted frog (Rana pretiosa) may not have occurred in California as specimens may have been mis-identified. We have retained this species in the predictive modeling, but readers are warned that this may be incorrect.
- The Black-collared lizard (Crotaphytus insularis) has been split into C. insularis and C. bicinctores, the Mojave black-collared lizard. We were unable to distinguish their individual ranges from the CWHR range maps and have modeled them as a single species.
- Subspecies of the Western aquatic garter snake (Thamnophis couchii) were recently elevated to full species status. Because range data is not yet provided in CWHR for the Santa Cruz garter snake (T. atratus) or the Giant garter snake (T. gigas), our modeling and analysis are for T. couchii as originally classified.
MethodsDistribution of vertebrates were modeled based on a six-step procedure that takes advantage of the known habitat preferences of species. First, criteria were developed to select the species to be included in gap analysis. Second, the distributional limits of each selected species was determined from available range maps. Third, the existing California Wildlife-Habitat Relationships (CWHR) database (Airola 1988) was used to assign suitability ratings to habitat types. Fourth, the land-cover map (Chapter 2) was reclassified as wildlife habitat types. Fifth, the range data and the CWHR database were used in a GIS-modeling process that assigned species to mapped habitat polygons (Chapter 2). Distributions were also summed for 7.5 minute quadrangles to create maps of species richness for each taxonomic group. Sixth, the modeling was compared to observed species lists for many parks and with the Breeding Bird Survey data.
There are 650 terrestrial vertebrate species included in the CWHR database. Some of these are introduced. Some do not breed in the state or are rare visitors. Some are marine mammals and pelagic birds that would not be appropriately modeled with GAP data on terrestrial environments. Therefore we shortened the list by the following criteria:
- Exclude introduced species such as the bullfrog (Rana catesbeiana).
- Exclude species not related to habitats of the terrestrial surface of the state, i.e., marine mammals, pelagic birds, waterfowl, bats.
- Exclude species that do not breed in the state, such as migratory birds that only winter in California.
Following this screening step, 455 species remained, including 205 birds, 134 mammals, 45 amphibians, and 71 reptiles.
The biogeographic range of each species was represented as presence or absence in 7.5 minute quadrangles based on digital maps of range limits that were produced by the California Department of Fish and Game for the three-volume series on California wildlife (Zeiner et al. 1990). These maps were originally prepared at 1:3,500,000 scale based on available scientific literature, museum records, and major physical and/or vegetational features. Some of these maps have been more recently revised at a 1:1,000,000 scale by Fish and Game. The range maps were summarized as a look-up table by Fish and Game as a list of quadrangle-species pairs. The November, 1997, version of the ranges by quadrangle look-up table was used for our predictive modeling. The exception to this was for birds, where the quadrangle lists did not distinguish breeding, summer range from winter range for migratory birds. For the birds, therefore, we compiled new lists by quadrangle from the latest set of range maps.
The suitability ratings by habitat type for each species were obtained from Version 5.3/6.0 of the California Wildlife-Habitat Relationships database. This database had been compiled and revised by an interagency team of wildlife biologists to contain all available information on habitat requirements or terrestrial vertebrates (Airola 1988). CWHR ranks each habitat type as High, Medium, Low, or Unsuitable for breeding, feeding, and cover. For gap analysis, we only employed the suitability rating for breeding.
In Chapter 2, we described the development of the CA-GAP land-cover map. In addition to classifying landscape units into plant community types, we also classified them into CWHR habitat types. Barry Garrison, the Fish and Game manager of the WHR Program, assisted us in cross-walking the combination of dominant species in each polygon into the habitat types described in the CWHR system (Mayer and Laudenslayer 1988, Schultze 1994). As each polygon in the vegetation map is considered to be a landscape mosaic of several habitat types, three major CWHR vegetation types can be assigned to each polygon as well as several different CWHR wetland/riparian types, the latter being coded as attributes of each polygon during the original mapping. The cross-walking procedure used a scoring system that rated the propensities of plant species in the CA-GAP land-cover database to be associated with various CWHR habitat types. The habitat type with the highest combined score for the combination of plant species was assigned to the polygon.
A species can then be predicted to be present or absent in a landscape map unit through the following GIS-based modeling process. The map of habitat types is overlaid with quadrangles. For each polygon, the model refers to a look-up table to determine if the polygon falls within the species' range as represented by the quadrangle maps. If so, the model then checks another look-up table of habitat suitability rankings for each of the three habitat types recorded in the polygon. Then the areal extent of each suitability rank in every habitat polygon is summed, accounting for the relative proportion of the primary, secondary, and tertiary habitat types present. Thus a habitat polygon might be modeled as having 30% High suitability habitat, 10% as Medium, 20% as Low and the remaining 40% as unsuitable. The model may also indicate that one or more of the wetland/riparian habitats originally recorded as polygon attributes may also be suitable (or in fact, only the wetland/riparian habitat may be suitable). Rather than simple presence/absence results, therefore, the CA-GAP wildlife modeling produced a vector of proportions of suitability levels for each polygon.
This modeling process can create problems at the edge of a species range limits. If any part of the habitat polygon overlaps a quadrangle within the range, one option would be to model the entire habitat polygon as potentially suitable habitat. For large habitat polygons spanning many quadrangles, this alternative solution could predict species presence far beyond its actual range limit. Alternatively, the predicted distribution could be arbitrarily truncated at the quadrangle boundary, creating an unnatural, rectilinear edge. Our solution was to check the proportion of each polygon within the range limits. If less than a majority of the habitat polygon was within the range limits as summarized by quadrangle, the entire habitat polygon was considered unsuitable.
To visualize these complex, multivariate data, we categorized the output into a combination of area and suitability to give a single class per polygon that could be displayed and analyzed further (Table 3-1). Thus a polygon that contained a large proportion of the best habitat could be distinguished from one with only a small amount or one where an arbitrary threshold for habitat suitability is used to include or exclude polygons. We believe this categorization is more meaningful than simple presence/absence data in which all the suitability levels and their extent is ignored and avoids making arbitrary decisions in the predictive modeling phase about what suitability level or areal proportion should be used as a thresholds to include or exclude polygons from the predicted distribution. Note that the categories are assigned in descending order, so that a polygon is assigned to category 4 only if it does not also qualify in category 5. Category 1 is primarily restricted to those habitat polygons in which none of the three major habitat components of the landscape mosaic (i.e., primary, secondary, tertiary) are considered suitable for a species, but where one or more of the wetland/riparian types is suitable. These types were only recorded as being present in a polygon where their areal extent was too small to include them as one of the dominant types. This category is also used in some cases where Joshua trees are known to be present. If their density was unknown, as described in Chapter 2, we could not confidently assign the polygon to Joshua Tree Woodland habitat. Instead, the polygon was rated as Category 1. Consequently there is no associated areal extent for these minor habitat types and no way to determine in which higher suitability category they might be included. Category 1 may in fact be very critical habitat for the species, but we can not use that information in spatial analysis. If a wetland or riparian type was large enough to be one of the three major types in a polygon, however, that would be a factor in assigning the polygon to categories 2-5. For mapping species richness (below) and for gap analysis (Chapter 6), we only include suitability ranks of 4 and 5 and omit the lower suitability levels.
Criteria
Category
>50% High Suitability
5
>50% Medium or High Medium or High Suitability
4
>50% Low, Medium or High Suitability
3
<50% Low, Medium or High Suitability but >0%
2
Suitable habitat in wetland/riparian types only (no areal estimate)
1
No suitable habitat
0
Table 3-1. Categories of predicted habitat quality based on suitability and areal extent.
Once a list of species is ascribed to each polygon, the predicted distributions were summed into species richness maps for the four major taxonomic groups: birds, mammals, amphibians, and reptiles. This involved resampling the species distributions mapped on habitat polygons to a uniform grid system of equal-area units (Stoms 1994). We selected the 7.5 minute USGS quadrangles and counted every species present in any polygon within the quadrangle with a suitability category of 4 or 5. That is, only species with the best habitat quality polygons were counted. This count would be similar to what could be calculated directly from the quadrangle by species look-up tables used as input to the GIS-based modeling. The difference here is that habitat distribution and suitability has been used to refine the coarser level representation of species range.
The data on species by quadrangle also allowed us to examine the turnover of species between sites, that is the similarity of species in one quadrangle to the set of species in every other quadrangle. This information is complementary to species richness because it can illustrate that two equally rich sites may, in fact, have distinctly different species composition and one site could not be substituted for another solely on the basis of richness. Our measure of similarity is the Jaccard similarity coefficient, which is simply the percentage of species common to both sites in relation to their combined set of species. An index of 100% would indicate identical species composition and therefore the sites would be interchangeable in any conservation plan based on this biological information alone. A low index occurs when the two sites share very few species. The index is calculated as a "distance" between every pair of sites, and so a map of the similarity index must always be in reference to a specific quadrangle.
Results
Predicted Species Distributions
In this section, we present a sample of representative maps of predicted species distributions showing the relative suitability rank of each habitat polygon. (The full set of distributions can be viewed in the CD-ROM and GIS database products.) The complete table of total mapped distribution (suitability ranks 4 and 5 only) and percent area of the entire state of California, is included in the gap analysis table in Appendix 6-1.
Land birds in California tend to have relatively wide distributions. The ext ent of high and moderate suitability habitat predicted for most species was between 10-50%. Very few species had more than 50%, e.g., turkey vulture. The Black-headed grosbeak (Figure 3-1) had high and moderate suitability habitat predicted for 31% of California's land area. The highest quality habitats are found in the foothills surrounding the Great Central Valley, portions of the north coast and urban areas such as Sacramento and Los Angeles. (See the Limitations and Discussion section at the end of this chapter about the suitability of urban lands as species habitat).
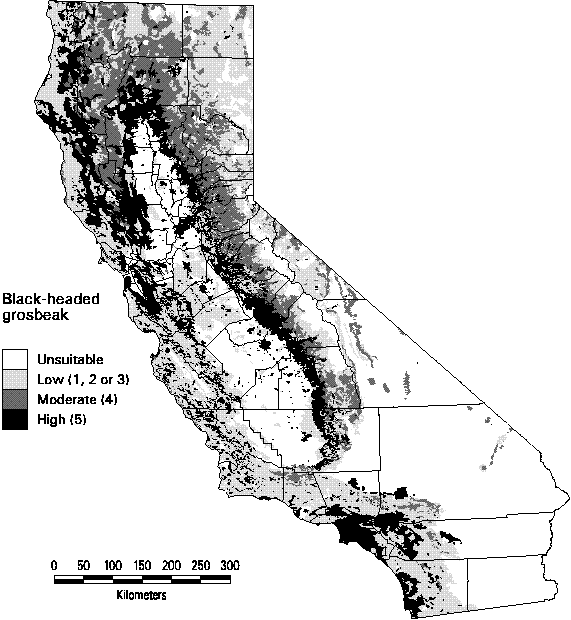
Figure 3-1. Predicted distribution and suitability categories of the Black-headed grosbeak (Pheucticus melanocephalus).
Several of the mammals were predicted to have high and moderate quality habitat over much of the land area of California. These species include the Black-tailed jack rabbit, Botta's pocket gopher, Western harvest mouse, deer mouse, coyote, american badger, and bobcat, all with distributions greater than 70% of the state. The Western gray squirrel is widespread (26%) through the montane areas of the state (Figure 3-2). Its highest suitability habitat, however, occurs in the area surrounding the Great Central Valley. Habitat suitability tends to be low on the central and southern coastal regions.
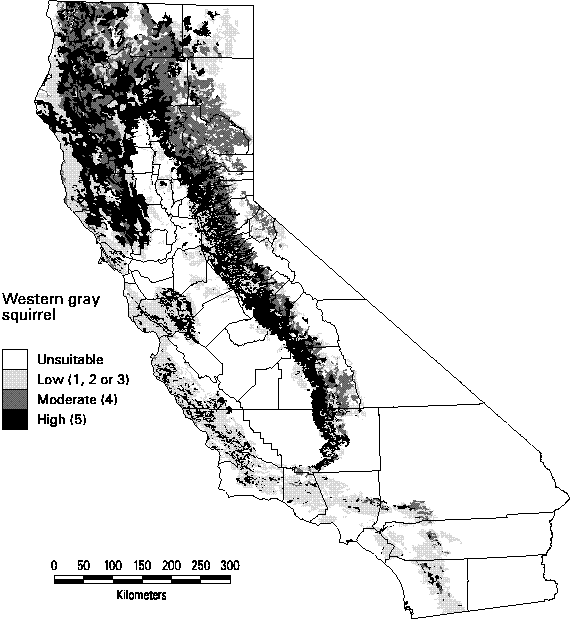
Figure 3-2. Predicted distribution and suitability categories of the Western gray squirrel (Sciurus griseus).
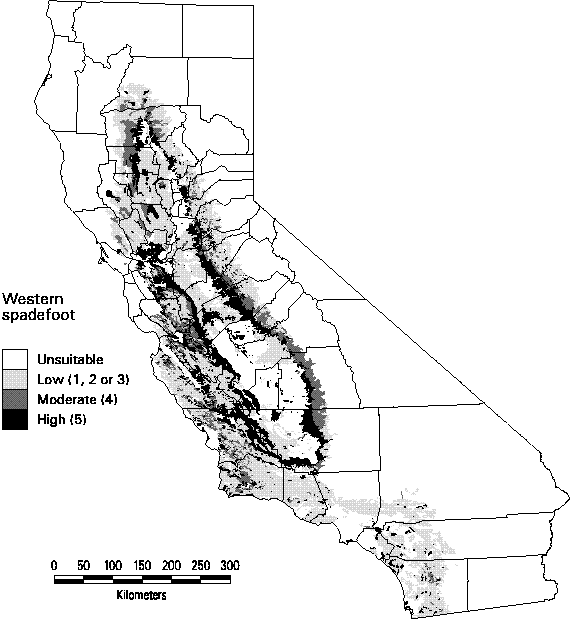
Figure 3-3. Predicted distribution and suitability categories of Western spadefoot (Scaphiopus hammondii).
Many of the amphibians have very narrow ranges in California, generally less than 10%. The widest predicted occurrence for any amphibian was for the Pacific chorus frog (Pseudaris regilla, WHR code # A039) with highly suitable habitat in nearly half the land area of the state. Several species were predicted to occur in no quadrangles with a suitability rank of at least 4. These species all had very small ranges, such as the Black toad (Bufo exsul) which only occurs in a single wetland in Deep Springs Valley in Inyo County. When the only suitable habitat for the species was wetland or riparian and this habitat was below the mapping resolution of CA-GAP, our method predicted a suitability rank of 1, which was not included in calculating species richness or in the gap analysis of predicted species distributions. The predicted distribution of the Western spadefoot is shown in Figure 3-3 which includes all suitability ranks. Although its range extends across the Great Central Valley and the central and southern coastal regions of the state, its most suitable habitat is relatively limited (roughly 11% of the state) to a narrow band around the valley.
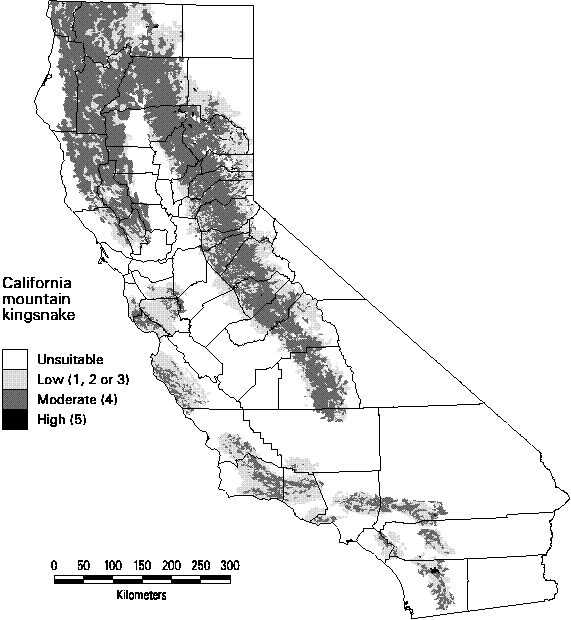
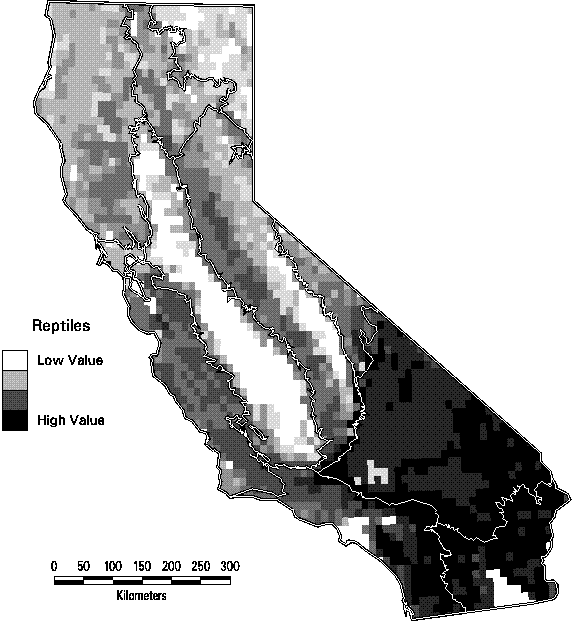
Reptiles tend to have wider distributions than the amphibians. Many species' distributions were predicted to exceed 20% of the state. With the exception of the gopher snake and the common kingsnake, however, no reptiles had highly suitable habitat over more the 50% of the state. The predicted distribution shown in Figure 3-4 is for the California mountain kingsnake. Although it is relatively widespread, there is virtually no high quality habitat (rank = 5) for this species.
Figure 3-4. Predicted distribution and suitability categories of California mountain kingsnake (Lampropeltis zonata).
Species Richness Patterns
Richness was determined by tallying all vertebrates with at least one polygon with a suitability rank of 4 or 5 in a quadrangle. Thus the entire quadrangle should not be assumed to be high suitability, only that some Medium or High quality is present. Similarly, richness is based on predicted habitat suitability, not on known occurrences of species. So richness itself is only an estimate of the actual number of species present. Because richness patterns are seldom the same among taxonomic groups (Prendergast et al. 1993), we computed it separately for the four orders of terrestrial vertebrates (Figures 3-5 through 3-8).
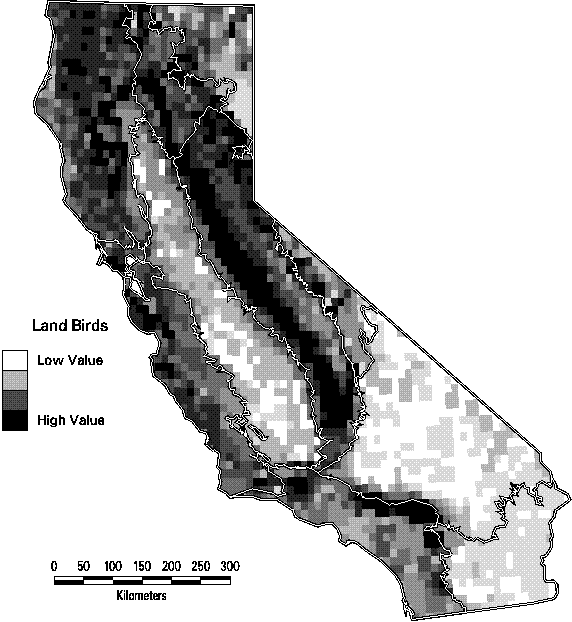
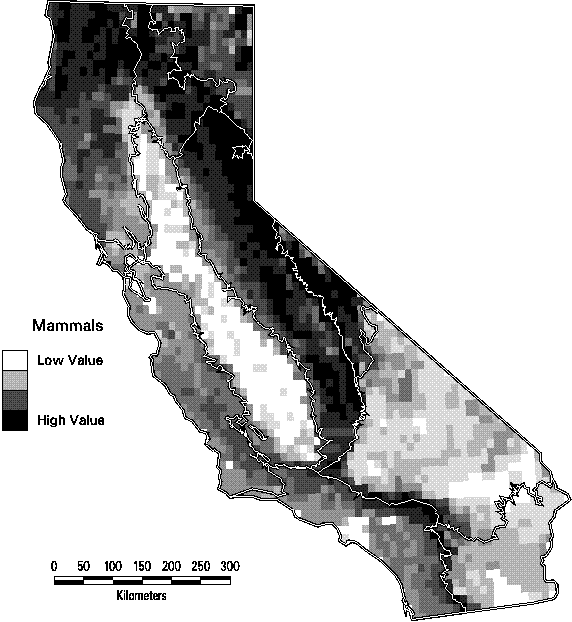
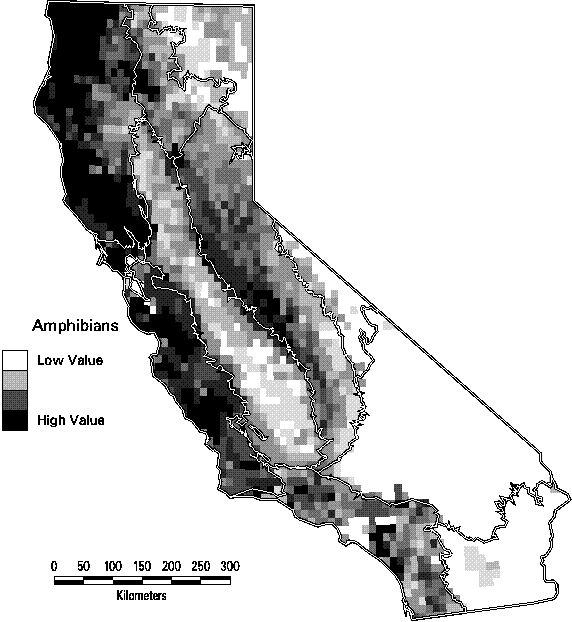
The patterns of species richness at the scale of 7.5 minute quadrangles are quite distinct among the four taxonomic groups. Land birds (summer ranges of breeding birds only) are most diverse in the lower elevations of the Sierra Nevada and Cascade ranges and in the mountains of southern California. The maximum number of species in a quadrangle is 126, or 61% of all species of land birds. Mammals have a similar pattern except that the richest quadrangles tend to be at higher elevations than birds. They also tend to have greater turnover in composition than birds, with a maximum of only 49% (66 species) of all mammals in the richest quadrangle. Amphibians are most diverse in the cooler, wetter parts of California, primarily on the northern and central Coast Ranges and the central Sierran foothills. Of the 45 native amphibians in the state, as many as 15 (33%) are predicted with highly suitable habitat in the richest sites , while many quadrangles in the deserts contain none. Reptiles, in contrast, thrive in the hot, arid deserts of southeastern California. Because of their concentration in just a portion of the state, the richest quadrangle for reptiles contains 73% (52 species) of all reptiles. Clearly, it would be impractical to base a conservation strategy solely on protecting the richest sites for one of the se groups, since it would seldom be a site that was rich for other taxa. If all terrestrial vertebrates are combined, the number of birds dominates the totals that the pattern is essentially the same as seen in Figure 3-5. As a result, the richest sites for all vertebrates, in the northeastern part of the state would capture few amphibians and reptiles.
Figure 3-5. Species richness of land birds (summer ranges of breeding birds only) by 7.5 minute quadrangle. (Only species with at least category 4 or 5 suitability are counted).
Figure 3-6. Species richness of mammals by 7.5 minute quadrangle. (Only species with at least category 4 or 5 suitability are counted).
Figure 3-7. Species richness of amphibians by 7.5 minute quadrangle. (Only species with at least category 4 or 5 suitability are counted).
Figure 3-8. Species richness of reptiles by 7.5 minute quadrangle. (Only species with at least category 4 or 5 suitability are counted).
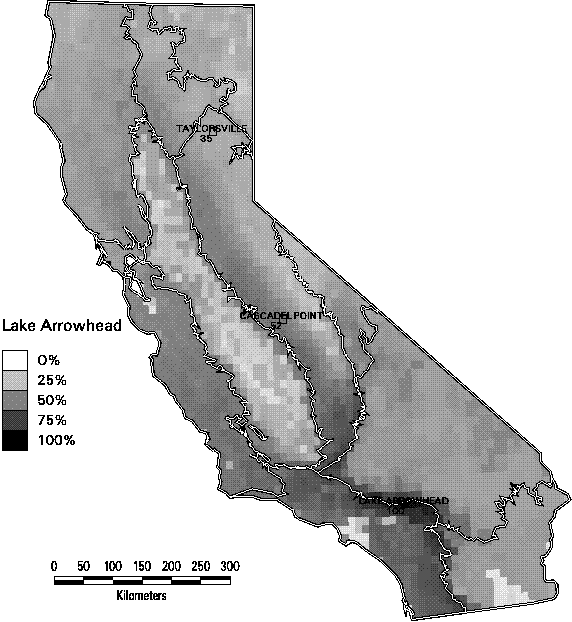
Richness alone can be a misleading indicator of biodiversity. In Figures 3-9 through 3-11, we illustrate one of the dilemmas of relying on species richness as a conservation criterion. Each of these 3 maps depicts the similarity in species composition of mammals of all quadrangles in relation to a reference quadrangle. For this set of maps, we selected 3 reference quadrangles that each contain 52 mammals (top 5% of all quadrangles in mammal richness), Taylorsville in the northern end of the Sierra Nevada range, Cascadel Point in the low- to mid-elevations of the southern Sierra, and Lake Arrowhead in the San Bernardino Mountains in southern California. All 3 quadrangles are characterized by a mix of montane conifer and hardwood forest habitats with some shrub types. Yet they show little similarity in composition. The two Sierran sites only share 2/3 of their combined species. The Taylorsville and Lake Arrowhead quadrangles share only 1/3. In fact, it is striking how rapidly the similarity of species composition declines as one moves away from the reference quadrangle. The average similarity of all other quadrangles with these 3 ranged from only 36-42% Selecting one site to represent biodiversity, simply because the site has high species richness, would tend to miss many species. This result emphasizes the importance of the complementarity of sites, since each contains species not represented by the others.
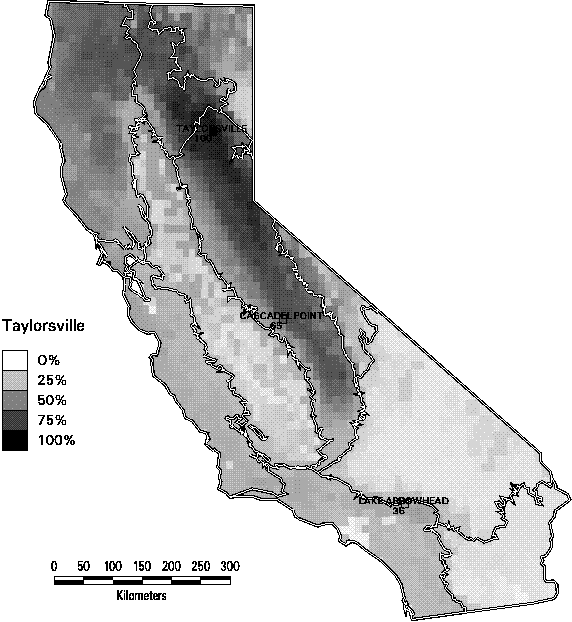
Figure 3-9. Jaccard similarity index of mammals by 7.5 minute quadrangle in reference to the Taylorsville quadrangle in the northern Sierra Nevada region. (Only species with at least category 4 or 5 suitability are counted).
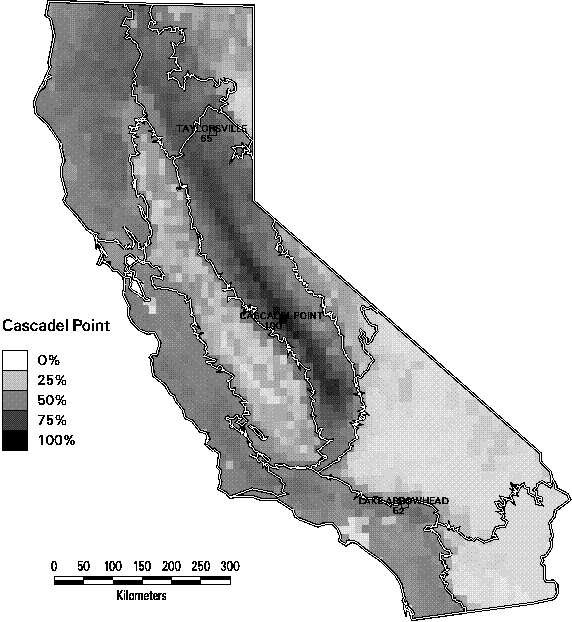
Figure 3-10. Jaccard similarity index of mammals by 7.5 minute quadrangle in reference to the Cascadel Point quadrangle in the southern Sierra Nevada region. (Only species with at least category 4 or 5 suitability are counted).
Figure 3-11. Jaccard similarity index of mammals by 7.5 minute quadrangle in reference to the Lake Arrowhead quadrangle in the Southwestern California region. (Only species with at least category 4 or 5 suitability are counted).
Accuracy Assessment
When this report was written, was written, the accuracy assessment/validation of the predicted species distribution modeling had not been completed. Two approaches are being taken concurrently: 1) comparison of predicted species lists using the CA-GAP data and methodology with lists of observed species from state parks and other manages areas (in collaboration with Dr. James Quinn at University of California, Davis), and 2) comparison of predicted species with the Breeding Bird Survey (BBS) data. Dr. Quinn's group at Davis has assembled and edited species lists from a large number of managed areas. From the comparisons with his data, we hope to be able to estimated the overall quality of the CA-GAP vertebrate data and identify which groups of species tend to be predicted with greater or lesser accuracy. Because the managed areas are various sizes, we should be able to estimate the spatial resolution at which the predictions are of satisfactory accuracy. The expectation is that the predictions will be less accurate for small sites than for large ones, where many of the small local errors are overcome as one encounters more habitat types. For instance, predicted lists for the entire state should be virtually perfect. All species known to occur would be predicted to occur somewhere in the state. But in very small managed areas of 100 ha or less, generally the habitat suitability method tends to greatly overestimate the number of species (errors of commission). At the same time, the generalization of the CA-GAP land-cover map will often miss individual patches of unique habitat and therefore fail to predict species at the site (errors of omission). The difficulty in making these comparisons is standardizing the predicted and observed lists to include the same types of species. For instance, park species lists often include every species ever seen in the managed area, not just the breeding species modeled in gap analysis.
The BBS is a long-term program of the U. S. Fish and Wildlife Service to collect data on the trends in birds across the nation. Each spring, 25 mile long sections of a sample of roads are surveyed, with stops at 0.5 mile intervals. Birds that are seen or heard are recorded, both the species and the number of individuals, in a brief stay at each stop. Comparisons of the predicted bird lists and the BBS will allow us both to test the CA-GAP wildlife predictions with actual field data over a large area as well as to evaluate the significance of management status of the transects in predicting the trends in bird abundance. If the assumptions of gap analysis are valid, we should expect to see declines in the birds that are least well-represented in managed areas.
Limitations and DiscussionThe methods followed by CA-GAP made use of the best available information to create meso-scale maps of predicted species distribution, but did not involve the collection of new specimens or field observations. Because locational records of collection and observation of species are grossly incomplete, CA-GAP relied strongly on the use of expert opinion in the development of range limits, habitat association, and review of predicted distributions. The resulting maps then are testable hypotheses that we encourage field biologists to assess whenever conducting field studies.
These procedures work best for species with habitat preferences that can be described in terms of land cover and other digitally mapped features or characteristics. It provides less accurate predictions of the presence of species that are highly variable in spatial and/or temporal occurrence (Krohn 1996), and it will work for habitat specialists only if their specific habitat requirements are available as mapped features or are well associated with other mapped characteristics such as land cover types.
An additional caution is that species with very restricted distributions (occurring in one or a few locations of a size below the GAP MMU) cannot be predicted to occur in seemingly appropriate habitat within their distributional limits. Because of their rarity, these species are often the subject of special attention from state and federal resource agencies and are ranked G1-G2 by the Natural Heritage Central Databases (see ranks and their description at http://www.heritage.tnc.org). The specific locations where they are known to occur are usually tracked by NHPs, CDCs, and the USFWS. GAP makes use of the data from NHPs and CDCs to report the presence of populations of such species within a geographic unit. For security purposes, the exact locations of these populations should be distributed only by the data owner.
It was noted earlier that urban habitats were rated in CWHR as High or Medium suitability for some species, such as the Black-headed grosbeak. CWHR does not distinguish between land use types within urban areas as it does for agricultural types and so the ratings are applied equally to all habitats from low density housing to fully developed industrial areas. Neither the CA-GAP land-cover nor the CWHR suitability databases have sufficient detail to discriminate between these different levels of use. We examined the CWHR habitat suitability ratings to determine the extent this would misrepresent both the distribution of some species but also their management status. Many of the species with this habitat rating are non-natives, which had been eliminated from our analysis. Of the remaining native species, perhaps 10% had this type of rating. Most of these were common birds that one would expect to be found at least in suburban, residential areas. In general, the species potentially affected by this simplification are not considered vulnerable.
Successful assessment of the management status of vertebrate species through gap analysis requires accurate mapping of their distributions. As of the date of this report, the accuracy assessment in still in progress, so quantitative measures of accuracy are not yet available.
There have been many concerns expressed about species richness criteria in conservation assessment and planning. Our purpose is not to identify conservation priorities as to portray relatively fine-grain biogeographic information that has not been available until compiled by CA-GAP. Species richness patterns are highly dependent on the size of the sampling unit (Stoms 1994). Our portrayal of species richness by 7.5 minute quadrangle is just one of many possible representations at other scales. We also constrained the count of species to be those with the highest categories of suitable habitat (i.e., 4 or 5). Consequently, the richness maps ignore species with small areas of highly suitable habitat or larger areas of low suitability habitat that may actually contain such species.
CA-GAP Home | Overview | Report | Download GIS | CD-ROM | Site Index | National GAPTop of Page Send your comments to: stoms@geog.ucsb.edu
UCSB Biogeography Lab Home